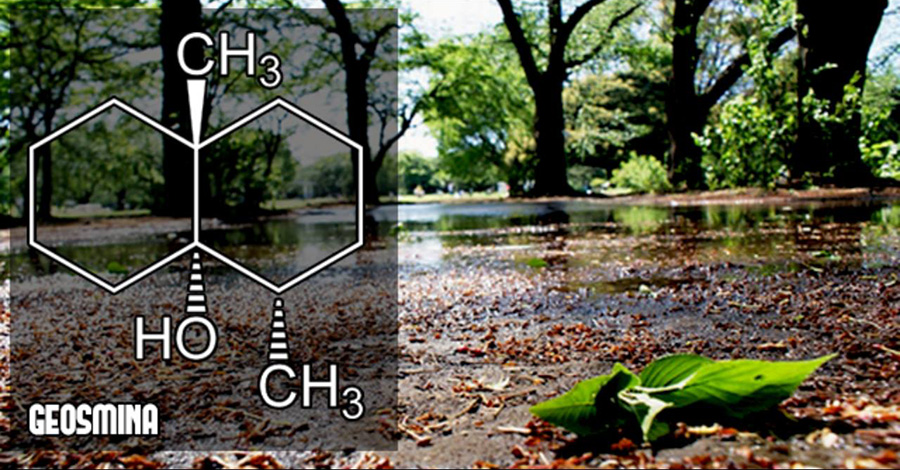
Tomado y ampliado a partir de ABC y la publicación original Cuando llueve las primeras gotas que caen sobre un suelo seco favorecen que se volatilice la geosmina, un compuesto producido por bacterias del género Streptomyces que viven en el suelo, y creando un aroma que tiene el evocador nombre de petricor o «tierra mojada».
Imagen tomada de https://tiwel.es/geosmina-olor-a-tierra-mojada/
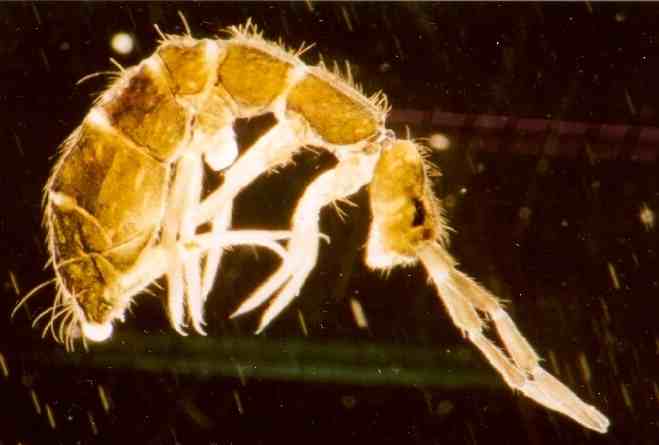
¿Por qué es agradable oler una molécula producida por microorganismos del suelo? Una investigación publicada en «Nature Microbiology» por el grupo de Klas Flardh esta semana ha podido responder, en parte, a esa pregunta. Después de colocar pequeñas trampas en el campo y de hacer experimentos en el laboratorio, han descubierto que la geosmina resulta atractiva para los colémbolos, unos minúsculos artrópodos saltarines muy abundantes en el suelo. Y podrían no ser los únicos artrópodos atraídos por este aroma.
Los colémbolos (Collembola) son un orden de artrópodos hexápodos cercana a los insectos, y a veces se les clasifica dentro de ellos. Son animales diminutos (3,5 mm), ubicuos, que ocupan todos los continentes (incluso la Antártida). Son, probablemente, los animales más numerosos de la Tierra: hasta 62.000 individuos por m2. Se conocen casi 7.900 especies y su registro fósil data desde el Devónico (Rhyniella, Protoisotoma, hace unos 400 millones de años) siendo, pues, uno de los animales terrestres más antiguos.
Imagen tomada de Wikipedia
Klas Flardh y colaboradores descubrieron que los colémbolos son grandes admiradores de la geosmina; que pueden percibirla con sus antenas y que acuden a su origen por un motivo muy concreto: alimentarse de las bacterias que liberan este olor. Pero los autores han averiguado algo más: la bacteria probablemente también saca provecho de esta irresistible atracción. «Cuando un colémbolo se alimenta de las colonias de Streptomyces, disemina sus esporas a través de las heces y por adherencia a su cutícula», explican los autores. Por tanto, se podría decir que todo esto es como una versión de la atracción de las abejas por el aroma de las flores pero en versión miniatura. «Los resultados indican que la producción de geosmina (...) es una parte integral del proceso de esporulación, fundamental para completar el ciclo de vida de Streptomyces al facilitar la dispersión de sus esporas por medio de los artrópodos», han concluido los investigadores.
Ciclo de vida de Streptomyces Publicaciones de Klas Flardh
Klas en un experto en la Biología Molecular de Streptomyces. Hizo la tesis con el Dr. Miguel Vicente en el CNB (Madrid) y Postdoct en el John Innes Center (Norwich, UK) con los que sigue manteniendo una estrecha relación. Tenemos un par de publicaciones conjuntas sobre el gen divIVA de Streptomyces y Corynebacterium glutamicum.
En todos los periódicos se ha publicado la noticia de que la ivermectina (descubierta por el premio Nobel Satosi Omura) se puede usar "in vitro"para reducir la carga viral del COVID-19. La secuenciación del genoma del microorganismo productor (Streptomyces avermitilis) se llevó a cabo en el laboratorio de Omura por Haruo Ikeda, compañero mío en el laboratorio de David Hopwood. Les adjunto el link al trabajo todavía no publicado. Las principales conclusiones son: Highlights
• Ivermectin is an inhibitor of the COVID-19 causative virus (SARS-CoV-2) in vitro.
• A single treatment able to effect ~5000-fold reduction in virus at 48h in cell culture.
• Ivermectin is FDA-approved for parasitic infections, and therefore has a potential for
repurposing.
• Ivermectin is widely available, due to its inclusion on the WHO model list of essential
medicines.
G. Schematic of ivermectin’s proposed antiviral action on coronavirus. IMPα/β1 binds to the coronavirus cargo protein in the cytoplasm (top) and translocates it through the nuclear pore complex (NPC) into the nucleus where the complex falls apart and the viral cargo can reduce the host cell’s antiviral response, leading to enhanced infection. Ivermectin binds to and destabilises the Impα/β1 heterodimer thereby preventing Impα/β1 from binding to the viral protein (bottom) and preventing it from entering the nucleus. This likely results in reduced inhibition of the antiviral responses, leading to a normal, more efficient antiviral response.
La palabra «mutación» tiene unas connotaciones realmente inquietantes en el imaginario colectivo. Los superhéroes de Marvel se transforman y adquieren capacidades extraordinarias gracias a las mutaciones y en más de una película de serie B una mutación hace de un escualo o a una piraña corriente una perversa máquina de matar. En la película « Estallido», que fue asesorada por el reputado epidemiólogo W. Ian Lipkin, una mutación hace que el ébola comience a transmitirse por vía aérea y se convierta en un patógeno que solo se puede detener con una bomba nuclear.
La pandemia de la COVID-19 ha superado al argumento de cualquier película de catástrofes. Es, según los científicos Yong-Zhen Zhang y Edward C. Holmes, «la tormenta epidemiológica perfecta». El protagonista de esta historia es un virus minúsculo e insignificante, de nombre SARS-CoV-2, que parece proceder de murciélagos y al que la evolución le ha permitido ser muy contagioso entre personas y causarles neumonía. Esta «película» no se habría desarrollado si no fuera porque el virus apareció en el lugar perfecto (Wuhan, China, una gran ciudad y un nudo de comunicaciones) y en el momento perfecto (antes del Festival de Primavera). De esta manera, una entidad replicadora no viva, que no entiende de fronteras ni de rivalidades políticas, ha puesto contra las cuerdas a medio planeta en solo unas semanas. Mientras la civilización se repone y se prepara para devolver el golpe, es legítimo preguntarse si las mutaciones pueden hacer que este coronavirus se transforme y se haga más letal o más escurridizo ante una vacuna.
Las mutaciones son errores de copiado
El virólogo Nathan D. Grubaugh, investigador en la Escuela de Medicina de Yale (EE.UU.), publicó en febrero un artículo titulado «No deberíamos preocuparnos cuando un virus muta en una epidemia», en la revista « Nature Microbiology». En este artículo explica que las mutaciones raramente tienen un efecto importante en una epidemia y define estos cambios como «Una consecuencia inevitable de ser un virus». Esto suele ser especialmente cierto en el caso de virus cuyo material genético es el ARN (no el ADN, como los animales o las plantas), tal como pasa con el SARS-CoV-2.
«La mutación es un aspecto rutinario de la vida para un virus de ARN», escribe Grubaugh. Éstos son especialmente negligentes a la hora de hacer copias de su material genético, una operación necesaria para multiplicarse e infectar, por lo que van acumulando fallos, auténticas erratas en sus secuencias genéticas, a las que también podemos llamar mutaciones. Todo esto ocurre porque «emplean una polimerasa de ARN (una enzima que actúa como copiadora de material genético) intrínsecamente dada a cometer errores, con lo que sus genomas acumularán mutaciones en cada ciclo de copia», explica Grubaugh.
Los errores cometidos al copiar su material genético son una importante fuente de diversidad para los virus - JdeVelasco
Las nubes de mutantes que nos infectan
Estas mutaciones tienen de su lado la fuerza de los números. Mientras que un humano necesita décadas y mucho esfuerzo para reproducirse, un virus puede multiplicarse en cuestión de horas y además puede replicarse miles de veces en cada célula de una persona infectada: en consecuencia, un mismo paciente no está infectado por un único virus, sino por muchas poblaciones distintas de virus, cada una con sus propias mutaciones.
«Un virus es en realidad una nube de mutantes», explica en este sentido Ignacio López-Goñi, catedrático de Micriobología de la Universidad de Navarra y autor de Microbioblog. «Tanto que se habla de cuasiespecies víricas, en vez de especies, para hablar de una población que comparte una secuencia consenso pero formada por distintos mutantes». De hecho, en el caso de infecciones duraderas, como las provocadas por el VIH, una misma persona puede ser infectada por «oleadas» de virus diferentes a medida que evolucionan dentro de su organismo.
Los virus no cambian tan rápido
Está claro que los virus tienen una capacidad incomparable de acumular mutaciones, pero esto no quiere decir que cambien tanto; suelen hacer falta años para que las mutaciones afecten a rasgos importantes de los virus. Los motivos son variados: «La mayoría de las mutaciones tienen un impacto negativo sobre alguna función del virus y acaban eliminadas por la selección natural», escribe Grumbaugh, lo que implica que desaparecen de la población porque adquirirlas es una desventaja. También pueden aparecer mutaciones que cambien la virulencia o la capacidad de transmisión del virus, pero según este investigador, «no se extenderán con mucha frecuencia a no ser que sean ventajosas».
Además, muchos de los rasgos de los virus, como el modo de transmisión, dependen de múltiples genes, así que es más improbable que cambios puntuales tengan un efecto beneficioso. Para que aparezcan ventajas hace falta que acumulen numerosas mutaciones y, por eso, «es raro ver virus que han cambiado en tiempos evolutivos cortos a pesar de tener altas tasas de mutación», en palabras de Nathan D. Grubaugh.
Las mutaciones no suelen aumentar la letalidad
Para nuestra tranquilidad, difícilmente estos cambios harán a los virus más letales, puesto que «La alta virulencia puede reducir la transmisibilidad del virus si el hospedador está demasiado enfermo», según este investigador. Es decir, si un virus deja a una persona moribunda o directamente fallecida, es muy probable que el patógeno deje menos descendientes capaces de seguir infectando.
Por eso, «Se suele ver todo lo contrario», dice José Antonio López Guerrero, virólogo y profesor de la Universidad Autónoma de Madrid. «Con el tiempo, los virus tienden adaptarse al hospedador y van perdiendo virulencia, porque no les interesa destruir a su hospedador. Así pasó con el virus de la gripe española o con la gripe de 2009». Un estudio subido recientemente al servidor « medRxiv» concluía que lo más probable es que el virus pierda severidad y se convierta en un patógeno estacional, recurrente cada invierno.
Dos trabajadores realizan tareas de desinfección en Taiyuan, en la provincia china de Shanxi, en febrero - REUTERS
Por tanto, las diferentes tasas de letalidad que se están registrando en cada país «probablemente no están causadas por la existencia de cepas más o menos virulentas», según López Guerrero. «Este baile de cifras es artificial: depende de la profundidad de las pruebas de PCR y de diagnóstico (si no se conoce la mayoría de los casos, solo se cuentan los casos más graves, con lo que la letalidad parece más alta) y de factores sociales y sanitarios en cada país», prosigue.
¿Pueden las mutaciones afectar a una vacuna?
Dejando al margen la letalidad, a medida que el coronavirus se extiende, sigue evolucionando en cada célula, en cada persona y en cada país y va acumulando nuevas mutaciones. Por eso podría llegar un punto en que las vacunas dejasen de ser útiles para nuevas variantes del SARS-CoV-2.
«Indudablemente un virus que muta mucho podría dar problemas», indica José Antonio López. «Pero éste no parece ser el caso del SARS-CoV-2, porque es bastante estable». Además, tal como explica, no solo hay decenas de candidatos a vacunas (se está trabajando en 41 de ellas), sino que también se busca diseñarlas para reconocer varias «dianas» de los virus y para generar una respuesta inmunitaria prolongada. Ignacio López-Goñi incide en que este coronavirus es menos variable que el virus de la gripe, «un campeón de la variabilidad» para el que se desarrollan nuevas vacunas cada temporada.
Holmes y Zhang recuerdan que este virus cuenta con una enzima con capacidad de corregir errores, compensando así su tendencia natural a copiar negligentemente el material genético. Por tanto, no hay pruebas de que el virus pueda sufrir «cambios radicales, como su transmisibilidad o virulencia, puesto que esos raramente cambian en la escala de una sola epidemia», escriben en un artículo publicado en « Cell».
En este sentido coincide Trevor Bredford, epidemiólogo en el Centro de Investigación Fred Hutchinson y profesor en la Universidad de Washington (EE.UU.). «Mi predicción es que deberíamos ver mutaciones ocasionales en la proteína S del SARS-CoV-2 (crucial para su capacidad de infectar) que le permitan escapar parcialmente de las vacunas o de la inmunidad de grupo, pero éste es un proceso que seguramente llevará años y no meses», tal como escribió en Twitter.
El «árbol genealógico del virus»
Aunque las mutaciones del SARS-CoV-2 no vayan a cambiar la letalidad o a alterar la eficacia de las vacunas, a corto plazo, hay muchos científicos colaborando y aprovechándose de las mutaciones para estudiar su evolución. Ya se ha registrado la existencia de «clústers» o grupos de virus en los países con distintas secuencias y se han secuenciado cientos de genomas.
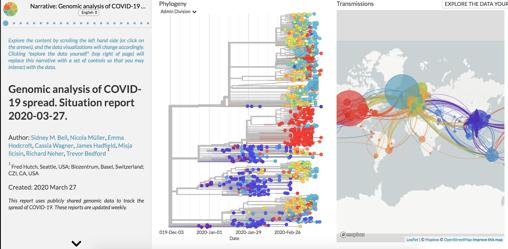
La última actualización de la web Nexstrain, promovida entre otros por Trevor Bredford, muestra el conocimiento recabado sobre la filogenia del coronavirus desde su salida de China: algo así como un árbol genealógico donde se puede observar la historia de los contagios. Ya se han secuenciado 1.495 genomas procedentes de 48 países en seis continentes. Los últimos datos muestran que el virus se ha estado diseminando y mezclando por Europa en las últimas tres a cinco semanas.
Captura de la web Nexstrain. Las mutaciones permiten reconstruir la historia de la transmisión del virus
Todo este trabajo permite estudiar, a nivel molecular, toda la historia del SARS-CoV-2: qué virus es, si es nuevo, cómo se transmite, cuál es su reservorio, qué factores provocaron su aparición, cómo evoluciona y cómo se adapta. Las técnicas de secuenciación de última generación, la publicación rápida de artículos y la colaboración internacional de científicos, compartiendo datos, permiten avanzar muy rápidamente en el conocimiento sobre el enemigo. Eso sí, para ser plenamente eficaces, estas técnicas tienen que combinarse con estudios epidemiológicos (que miden sobre el terreno cómo avanza la epidemia) y con análisis serológicos, masivos sondeos que rastrean la presencia de anticuerpos frente al virus para saber cuál es realmente la dispersión del virus.
¿Cómo son los mutantes del SARS-CoV-2?
En esta pandemia de récord la respuesta científica tambien ha sido de récord. Tan solo unos días después de que se detectase la epidemia, los científicos lograron una de las cosas más importantes en la respuesta ante estas situaciones: hacer el «retrato robot» del enemigo. El 5 de enero, gracias a las más avanzadas técnicas de secuenciación, científicos chinos obtuvieron el genoma completo del virus. Esto permitió averiguar quién era y de dónde venía, lo que es clave para comprender cómo se transmite o cómo infecta y para diseñar nuevos tratamientos y vacunas. De hecho, solo dos meses después de obtener su secuencia ya se están probando candidatos a vacunas, como el de la compañía Moderna, en pacientes voluntarios.
Representación del SARS-CoV-2 - CDC
Gracias a esta secuencia, hoy se sabe que el SARS-CoV-2 es un coronavirus muy similar al que provocó la epidemia de 2003 (ahora llamado SARS-CoV-1) y que pertenece al género de los Betacoronavirus. Sin embargo, se ha averiguado que, gracias a una larga historia de mutaciones y de evolución este virus tiene modificada una proteína de su envuelta, la S, que le permite unirse a las células a las que va a atacar, y que la secuencia genética de ésta es un 28% diferente a la del primer coronavirus. También se ha descubierto una modificación en la secuencia de la proteína S que le permite unirse a la furina, una enzima de animales. Estos cambios, aparte de otros, han convertido al SARS-CoV-2 en un virus altamente peligroso, por su capacidad de transmitirse entre personas.
El misterioso origen del coronavirus
Los análisis también han permitido mostrar que el SARS-CoV-2 es similar a coronavirus que infectan a murciélagos y pangolines, lo que lleva a sospechar que proceda de los primeros y que los segundos sean hospedadores intermedios.
La acumulación de mutaciones detectada, implica, según escriben Edward C. Holmes y Yong-Zhen Zhang en su artículo de «Cell», «más de 20 años de evolución de secuencias». Es decir, el virus no mutó repentinamente, sino que la actual pandemia se gestó durante décadas en varios animales sin que lo detectáramos. Otra posibilidad que no se puede descartar es que el virus evolucionase hasta su versión actual transmitiéndose durante años entre personas, sin que lo detectáramos hasta diciembre de 2019, según discuten Holmes y Zhang.
Un pangolín. El SARS-CoV-2 podría haber infectado a estos animales antes de saltar al humano
Además, el genoma del SARS-CoV-2 y su historial de mutaciones están lejos de indicar que proceda de un laboratorio, como ha sostenido alguna teoría. Su secuencia se publicó rápidamente y ha sido investigada por científicos de todo el mundo que, además, han ido obteniendo nuevas secuencias a medida que la epidemia se extendía. Un artículo publicado la semana pasada en « Nature Medicine» repasaba las pruebas de que el mutante SARS-CoV-2 no ha sido fabricado por el humano: fundamentalmente los cambios que presenta no tienen nada que ver con lo predicho por los modelos y no se usan los sistemas enzimáticos que se conocen. «Los datos genéticos revelan irrefutablemente que SARS-CoV-2 no proviene de ningún "chasis" de virus usado previamente», escriben los autores de ese estudio.
Con un tercio de la población mundial confinada en el interior de sus casas, el planeta entero parece haberse detenido de repente. Numerosos equipos de investigadores luchan contra el tiempo para poner a punto una vacuna mientras los sistemas sanitarios se colapsan y el coronavirus sigue avanzando y cobrándose nuevas víctimas cada día. ¿Cuándo podremos retomar nuestras vidas con normalidad? La respuesta no es fácil, y depende de una complicada serie de factores que, además, varían de una a otra región del planeta. El consenso de los científicos, sin embargo, parece unánime: incluso si el número de contagios empezara a disminuir en el transcurso de las próximas semanas, o meses, estaríamos aún muy lejos del final. Y el coronavirus tardará mucho tiempo en desaparecer, posiblemente años. La actual política de confinamiento es, sin duda, muy efectiva a la hora de frenar la curva de contagios, pero eso no implica que el virus vaya a desaparecer. La actual estrategia de cerrar amplios sectores de la sociedad no resulta sostenible a largo plazo, ya que causaría daños irreparables tanto en la sociedad como en la economía. Pero levantar las restricciones que frenan el avance del virus supondría un coste inasumible en vidas humanas. Ante esta situación, los países necesitan con urgencia una estrategia de salida que les permita romper este círculo vicioso y seguir adelante. Un plan que permita volver a la normalidad sin que ello suponga el sacrificio de miles de ciudadanos. Y eso constituye un enorme desafío, tanto científico como social. Tres formas de salir Según un informe publicado recientemente por la BBC, existen tres formas posibles de salir de este desastre, aunque ninguna de ellas es la panacea. Son éstas:
1.- Vacunas:más de un año de espera Según los números, si se pudiera vacunar por lo menos a un 60% de la población, el coronavirus dejaría extenderse como lo hace ahora. Pero hace solo unos días que las primeras personas recibieron vacunas experimentales en Estados Unidos y China, y eso (por lo menos en el caso de EE.UU.) solo después de que se permitiera a los investigadores saltarse el paso obligatorio de probar primero con animales. Aún así, y aunque la investigación avanza a una velocidad que no tiene precedentes, no existen aún garantías de que esas vacunas vayan a ser exitosas. Y si todo sale bien, las primeras unidades de esas vacunas aún tardarán entre 12 y 18 meses en empezar a ser distribuidas. Demasiado tiempo como para mantener los actuales niveles de restricción. 2.- Inmunidad natural: por lo menos dos años Según explica el informe de la BBC, la estrategia a corto plazo del confinamiento no erradica el virus, ya que tiene como objetivo reducir en lo posible el número de casos para evitar el colapso de los sistemas sanitarios. Una vez que se llegue al tan esperado "pico de infección" y la curva de contagios se vuelva descendente, se podría permitir que algunas de las medidas restrictivas se levantaran durante algún tiempo, dando un respiro a la sociedad hasta que las infecciones volvieran a aumentar e hicieran necesaria otra tanda de restricciones. Esta situación, repetida una y otra vez, podría llevar a que cada vez más ciudadanos se volvieran inmunes lo que, con el tiempo, permitiría volver a poner en marcha a una parte cada vez mayor de la sociedad. Pero ningún experto cree que el número necesario de personas inmunes pueda llegar a ser el suficiente hasta dentro de, por lo menos, dos años. Y queda abierta, además, la cuestión de cuánto tiempo podría durar esa inmunidad. Otros coronavirus ya nos han enseñado que las respuestas inmunes frente a ellos son muy débiles, lo que lleva a que una misma persona pueda contagiarse varias veces a lo largo de su vida. 3.- Cambiar el comportamiento de la sociedad La tercera opción para recuperar una cierta normalidad sería llevar a cabo cambios permanentes en nuestro comportamiento, tanto individual como colectivo, lo que nos permitiría mantener siempre bajas las tasas de transmisión. Hacerlo podría significar mantener "sine die" algunas de las medidas actuales, estando siempre pendientes de la aparición de nuevos brotes para, cada vez que sea necesario, volver a endurecer las medidas. También el desarrollo de medicamentos capaces de tratar con éxito una infección por Covid-19 podrían ayudar a las tres posibles estrategias, evitando o limitando la transmisión entre personas. A largo plazo, pues, la mejor forma de salir de esta situación es esperar a la vacuna. Los científicos están tratando de acelerar su desarrollo lo más posible. Mientras, no quedará más remedio que cambiar el modo en que vivimos y nos relacionamos. ¡El mundo debe cambiar, y debe hacerlo ya!. Tomado de ABC
The methylotrophic yeast Pichia pastoris is widely used in the manufacture of industrial enzymes and pharmaceuticals. Like most biotechnological production hosts, P. pastoris is heterotrophic and grows on organic feedstocks that have competing uses in the production of food and animal feed. In a step toward more sustainable industrial processes, we describe the conversion of P. pastoris into an autotroph that grows on CO2. By addition of eight heterologous genes and deletion of three native genes, we engineer the peroxisomal methanol-assimilation pathway of P. pastoris into a CO2-fixation pathway resembling the Calvin–Benson–Bassham cycle, the predominant natural CO2-fixation pathway. The resulting strain can grow continuously with CO2 as a sole carbon source at a µmax of 0.008 h−1. The specific growth rate was further improved to 0.018 h−1 by adaptive laboratory evolution. This engineered P. pastoris strain may promote sustainability by sequestering the greenhouse gas CO2, and by avoiding consumption of an organic feedstock with alternative uses in food production.
Fig. 1: Scheme for engineering chemoorganoautotrophy inP. pastoris.
Enlace al trabajo de Molecular Microbiology The twin‐arginine protein transport (Tat pathway) is found in prokaryotes and plant organelles and transports folded proteins across membranes. Targeting of substrates to the Tat system is mediated by the presence of an N‐terminal signal sequence containing a highly conserved twin‐arginine motif. The Tat machinery comprises membrane proteins from the TatA and TatC families. Assembly of the Tat translocon is dynamic and is triggered by the interaction of a Tat substrate with the Tat receptor complex. This review will summarises recent advances in our understanding of Tat transport, focusing in particular on the roles played by Tat signal peptides in protein targeting and translocation.
Fig. 1. Targeting to the Sec and Tat pathways. (a) The Sec pathway transports unfolded proteins. During co‐translational targeting to Sec, the signal sequence is recognised at the translating ribosome by ribosome‐bound signal recognition particle (SRP) and the nascent chain is guided via the SRP receptor to the Sec translocon, where the energy of protein synthesis is harnessed to drive protein transport. In the post‐translational pathway, the substrate is maintained in an unfolded conformation and guided to the Sec translocon by the ATPase, SecA. ATP hydrolysis by SecA provides the driving force for Sec‐dependent post‐translational protein export (Collinson et al., 2015; Lycklama a Nijeholt & Driessen, 2012; Rapoport, Li, & Park, 2017; Tsirigotaki et al., 2017). The Tat pathway transports folded proteins without the requirement for targeting factors. (b) Signal peptides that target to Sec and Tat pathways share a similar tripartite organisation with a positively charged n‐region, hydrophobic h‐region and polar c‐region containing a signal peptidase cleavage site (AxA). Tat signal peptides have an almost invariant pair of arginines that are embedded within a SRRxFLK motif (Berks, 1996). A helix destabilising residue (#), often a glycine, serine or proline towards the C‐terminal end of the h‐region, provides flexibility at this region of the signal peptide (Hamsanathan et al., 2017). A basic residue (+) is frequently found in the Tat signal peptide c‐region and serves as a Sec avoidance motif (Bogsch et al., 1997). The arrow indicates the position of signal peptide cleavage. Amino acid sequences of two E. coli Sec signal peptides, OmpA (post‐translational Sec targeting; Fekkes et al., 1998) and DsbA (co‐translational targeting; Schierle et al., 2003)—basic residues in the n‐region and the signal peptidase cleavage site in the c‐region are underlined and shown in bold. Two well‐studied E. coli Tat signal peptides, SufI and TorA, are also shown. Residues that match the twin‐arginine consensus are in red, the Sec avoidance signal in bold typeface and the signal peptidase cleavage site in underline
A Model for the Tat transport pathway. Step1. A folded Tat substrate docks at the Tat receptor complex, the twin‐arginines in the signal peptide n‐region binding to the cytoplasmic surface of TatC. Step 2. The signal peptide transitions to bind more deeply into the receptor, inserting in a hairpin conformation. The deep insertion of the signal peptide displaces TatB from its resting state binding site on TatC to occupy the TatA binding site at TMH6. A TatA molecule is now recruited to the binding site vacated by TatB. Step 3. The positioning of TatA at the TM5 binding site allows the further recruitment and nucleation of TatA molecules to form a large oligomer. Step 4. The signal peptide hairpin unhinges and the substrate passes across the membrane facilitated by the TatA oligomer. Step 5. The signal peptide is cleaved and the mature domain is released at the periplasmic side of the membrane. Following substrate translocation, the TatA oligomer dissociates and the Tat receptor returns to the resting state
In recent decades, the rapid expansion of antibiotic use and intake of high-calorie, low-fiber diets have contributed to disturbances in the gut microbial community, predisposing humans to various diseases such as metabolic syndrome. Although the influence of microbiota on the postnatal environment has been well documented, much less is known regarding the impact of gut microbiota at the embryonic stage. Although accumulating evidence supports the notion of the developmental origins of health and disease (DOHaD), the underlying mechanisms remain obscure. In this study, we explored the impact of maternal gut microbiota on embryonic development and disease susceptibility late in life.
RATIONALE
Gut microbiota–derived metabolites represented by short-chain fatty acids (SCFAs; e.g., acetate, propionate, and butyrate) not only fuel host cells but also serve as signaling molecules between the gut microbiota and extraintestinal organs. GPR41 and GPR43 belong to the free fatty acids receptor (FFAR) family and are receptors for SCFAs. We have previously corroborated the biological importance of FFARs in energy metabolism through interactions with dietary ingredients, as well as gut microbiota–derived metabolites. Gut microbial SCFAs regulate host energy homeostasis via GPR41 and GPR43 in the sympathetic nervous system, adipose tissues, pancreas, and intestine. A recent study further showed that the gut microbiota of pregnant mice influence immune and brain functions of offspring. These findings raise the possibility that maternal SCFAs play a key role in the regulation of disease susceptibility during postnatal life in the context of the DOHaD theory.
RESULTS
We found that maternal microbiota during pregnancy imparts resistance to obesity to their offspring. Pregnant mice were bred under specific pathogen–free (SPF) and germ-free (GF) conditions, after which newborns were raised by foster mothers under conventional conditions to align growth environments after birth. The offspring from GF mothers were highly susceptible to metabolic syndrome characterized by an exacerbation of obesity and glucose intolerance in association with reduced energy expenditure upon high-fat diet consumption during adulthood. A similar phenotype was also observed in offspring from mice fed a low-fiber (LFi) diet during pregnancy. Treatment of pregnant GF or LFi-fed mice with SCFA rendered adult offspring resistant to obesity. SCFA in the colonic lumen of pregnant mice reached the embryos via the maternal liver and bloodstream. Notably, the sympathetic nerves, intestinal epithelium, and pancreas of embryos highly expressed GPR41 and/or GPR43 to sense SCFAs originating from the maternal gut microbiota. Deficiency in embryonic GPR41 and GPR43 signaling compromised energy metabolism because of sympathetic dysfunction and hyperglycemia during the prenatal period. The SCFA-GPR41 and SCFA-GPR43 axes facilitate the development of neural cells, GLP-1–expressing enteroendocrine cells, and pancreatic β cells, thereby shaping embryonic energy metabolism. This developmental process contributes to maintaining postnatal energy homeostasis.
CONCLUSION
We determined that, during pregnancy, the maternal gut microbiota confers resistance to obesity in offspring via the SCFA-GPR41 and SCFA-GPR43 axes. During pregnancy, SCFAs from the maternal gut microbiota were sensed by GPR41 and GPR43 in the sympathetic nerve, intestinal tract, and pancreas of the embryo, influencing prenatal development of the metabolic and neural systems. These findings indicate that the maternal gut environment during pregnancy is a key contributor to metabolic programming of offspring to prevent metabolic syndrome. Thus, the gut microbiota of pregnant mice provides an environmental cue that fine-tunes energy homeostasis in offspring to prevent the developmental origin of metabolic syndrome.
During pregnancy, maternal gut microbiota influences offspring propensity for obesity via embryonic SCFA receptors.
The maternal gut microbial SCFA-embryonic GPR41 and GPR43 axes facilitate the development of neural cells, GLP-1–expressing enteroendocrine cells, and pancreatic β cells to shape the development of energy metabolism in offspring, even as adults. GPCR, G protein–coupled receptor.